
 Posted in
Posted in  Tags:
Tags: Следующий этап энергетического обмена, идущий за гликолизом, — клеточное дыхание, или, как его еще называют, биологическое окисление. Это кислородный этап окисления органических соединений. Если рассматривать дыхание в широком смысле слова, то это процесс поглощения живыми организмами кислорода (О2) из окружающей среды и выделения ими углекислого газа (СО2). Этот процесс необходим для поддержания внутриклеточных окислительных процессов, обеспечивающих энергетический обмен. Дыхание может быть внешним дыханием и тканевым или клеточным. Что такое внешнее дыхание понятно из названия. Так называют процесс газообмена между живым организмом и окружающей его средой. Тканевое или клеточное дыхание (еще называют биологическое окисление) – совокупность ферментативных окислительно-восстановительных реакций. В результате этих реакций сложные органические вещества окисляются кислородом до углекислого газа, при этом освобождается энергия, запасаемая клетками в форме АТФ.
Клеточное дыхание у растений, животных и большей части аэробных микроорганизмов начинается с отщепления СО2 (декарбоксилирования) от молекулы пировиноградной кислоты (пирувата), которая была образована в процессе гликолиза. Таким образом, гликолиз является необходимой подготовительной стадией клеточного дыхания при расщеплении углеводов. В процессе этой реакции от пирувата отрывается СО2 и образуется двухуглеродный остаток – радикал уксусной кислоты (ацетил-радикал). Этот двухуглеродный остаток присоединяется к молекуле универсального переносчика углеводородных радикалов — кофермента А — с образованием ацетил-кофермента А (ацетил-КоА). В результате этой реакции НАД+ восстанавливается до НАДН. Ацетил-КоА и НАДН образуются и при окислении жирных кислот, которые также являются субстратами клеточного дыхания. В дальнейшем окисление ацетил-КоА происходит в цикле Кребса, а НАДН – в дыхательной цепи митохондрий. В цикл Кребса на различных стадиях могут вступать все аминокислоты. Таким образом, в цикле Кребса сходятся пути окисления и углеводов, и жиров, и белков.
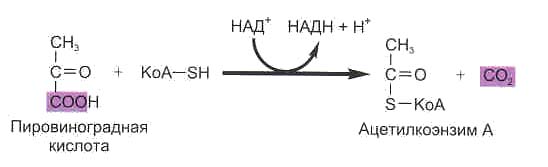
Отщепление молекулы углекислого газа от молекулы пировиноградной кислоты.
Цикл Кребса (также его называют цикл трикарбоновых кислот или цикл лимонной кислоты) – это сложный многоступенчатый окислительно-восстановительный процесс, в результате которого остаток уксусной кислоты, полученный от ацетил-КоА, полностью окисляется до 2-х молекул СО2 с образованием 3-х молекул НАДН, одной молекулы ФАДН2 и одной молекулы ГТФ. Все ферменты цикла Кребса также, как и ферменты окисления жирных кислот, локализованы в матриксе митохондрий, а один фермент – сукцинатдегидрогеназа – находится во внутренней митохондриальной мембране.
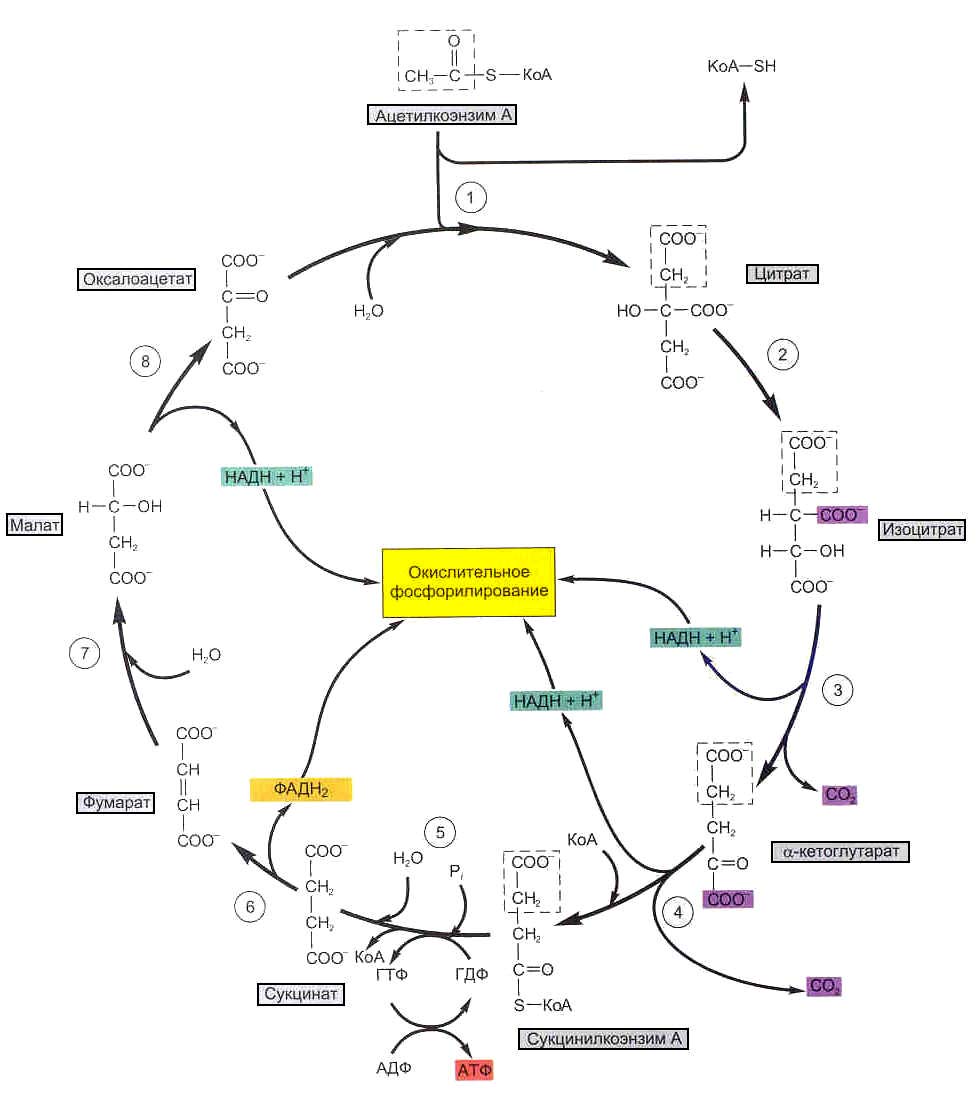
Цикл Кребса
На первой стадии цикла Кребса остаток уксусной кислоты передается от ацетил-КоА на молекулу щавелевоуксусной кислоты (оксалоацетата) с образованием лимонной кислоты (цитрата), которая через промежуточную реакцию образования цис-аконитовой кислоты превращается в изолимонную кислоту (изоцитрат). От изолимонной кислоты отщепляется СО2 и 2 атома Н+, в результате чего образуется молекула НАДН и a-кетоглутаровая кислота (a-кетоглутарат), которая взаимодействует с молекулой кофермента А. При этом отщепляется вторая молекула СО2 и образуется еще одна молекула НАДН и богатое энергией соединение сукцинил-КоА, которое расщепляется с образованием свободной янтарной кислоты (сукцината), что сопровождается синтезом ГТФ из ГДФ и Фн. Янтарная кислота окисляется до фумаровой (фумарата) с образованием ФАДН2, фумаровая кислота с присоединением воды превращается в яблочную (малат), а яблочная кислота окисляется до щавелевоуксусной (оксалоацетата) с образованием НАДН. На этой стадии цикл Кребса замыкается, т.е. оксалоацетат может снова вступать в цикл и конденсироваться со следующим остатком уксусной кислоты с образованием цитрата.
Таким образом, суммарную реакцию цикла Кребса можно описать следующим уравнением:
Ацетил-КоА +3НАД+ + ФАД + ГДФ + Фн +3Н2О —> 2СО2 + 3НАДН + 3Н+ + ФАДН2 + ГТФ + КоА
Энергия, освобождаемая при окислении ацетил-КоА, запасается в виде одной молекулы ГТФ (которая может превращаться в АТФ) и 4-х молекул восстановительных эквивалентов (3 молекулы НАДН и одна ФАДН2),
которые могут или использоваться в различных процессах биосинтеза, или окисляться. Дальнейшее их окисление происходит в дыхательной цепи митохондрий, которая локализована во внутренней митохондриальной мембране. При окислении НАДН в дыхательной цепи митохондрий происходит отрыв от него электронов, и их перенос на молекулу кислорода. У аэробных бактерий дыхательная цепь расположена в специальных структурах плазматической мембраны – мезосомах, и в общих чертах напоминает дыхательную цепь митохондрий.
| Входящий субстрат |
Ацетилкоэнзим А — источником ацетильной группы являются пируват, жирные кислоты и аминокислоты. |
| Локализация ферментов | Внутренние отделы митохондрий (матрикс) |
| Образование АТФ |
Непосредственно в цикле образуется одна молекула ГТФ, которая может быть превращена в АТФ. |
| Образование коферментов | 3НАДН + 3H+ и ФАДН2 |
| Конечные продукты |
Две молекулы CO2 на каждую молекулу ацетилкоэнзима А, входящую в цикл. Некоторые интермедиаты используются для синтеза аминокислот и других органических молекул, необходимых для осуществления функций клетки |
| Суммарная реакция | АцетилКоА + 3НАД+ + ФАД + ГДФ + Pi + 2H2O —> 2CO2 + KoA + 3НАДН + 3H+ + ФАДН2 + ГТФ |
Окислительное фосфорилирование начинается с окисления НАДН в дыхательной цепи митохондрий, сопровождающегося отщеплением двух электронов и протона (Н+). Окончательным акцептором этих электронов является О2, который соединяется с ионами Н+, находящимися в матриксе, с образованием Н2О. Электроны, отобранные от НАДН, передаются в дыхательной цепи от одного переносчика к другому, при этом они теряют свой восстановительный потенциал. Часть энергии, выделяемой при этом, рассеивается в виде тепла, но, кроме того, часть энергии тратится на создание на внутренней мембране митохондрий разности концентраций протонов (электрохимического потенциала) за счет их переноса в нескольких пунктах дыхательной цепи (так называемых пунктах сопряжения) из матрикса в межмембранное пространство.
Разность концентраций протонов получается в результате того, что при переносе электронов от НАДН к кислороду происходит «перекачивание» протонов из матрикса митохондрий в межмембранное пространство.
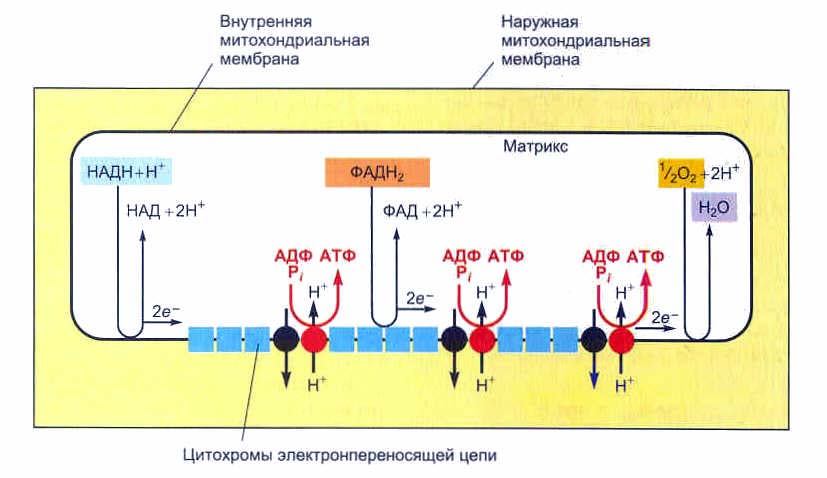
«Перекачивание» протонов из матрикса митохондрий в межмембранное пространство
В результате работы дыхательной цепи митохондрий концентрация Н+ в межмембранном пространстве намного выше их концентрации в матриксе, это создает направленный внутрь митохондрий градиент концентрации протонов. Мембрана митохондрий является для них непроницаемой, т.е. можно сказать, что она работает как плотина гидроэлектростанции, удерживающая воду в водохранилище. Энергия данного градиента используется ферментом АТФ-синтетазой, переносящим в матрикс ионы Н+ и синтезирующим АТФ из АДФ и Фн.
Для синтеза 1 молекулы АТФ необходимо перенести внутрь митохондрий 3 иона Н+ по градиенту концентрации, следовательно за счет окисления 1 молекулы НАДН может быть синтезировано 3 молекулы АТФ, а при окислении 1 молекулы ФАДН2 – 2 молекулы АТФ.
Кроме того, часть энергии градиента концентрации протонов тратится на перенос через внутреннюю мембрану митохондрий различных веществ. Синтез АТФ в митохондиях ферментом АТФ-синтетазой называют окислительным фосфорилированием, подчеркивая связь этого процесса с окислением органических субстратов.
| Входящие субстраты |
Атомы водорода, полученные от НАДН + Н+ и ФАДН2. |
| Локализация ферментов | Внутренняя мембрана митохондрий |
| Образование АТФ |
Три молекулы АТФ на каждую молекулу НАДН + Н+ |
| Конечный продукт | H2O — одна молекула на каждую пару водородов, входящих в цепь |
| Суммарная реакция |
1/4 O2 + НАДН + Н+ + 3АДФ + 3Pi —> H2O + НАД+ + 3АТФ |
Таким образом, в результате полного окисления глюкозы до углекислого газа CO2 и воды H2O образуется большое количество АТФ – 38 молекул. Две из них синтезируются в процессе гликолиза, а остальные 36 – при окислении пирувата. 1) при образовании одной молекулы пирувата в процессе гликолизе восстанавливается молекула НАДН, окисление которого в митохондриях дает 3 молекулы АТФ. 2) в процессе декарбоксилировании пирувата и образовании ацетил-КоА будет восстановлена еще 1 молекула НАДН (т.е. это 3 молекулы АТФ). 3) в цикле Кребса образуются 3 молекулы НАДН (это будет 9 молекул АТФ), 1 молекула ФАДН2 (плюс еще 2 молекулы АТФ) и 1 молекула ГТФ (обменивается своим терминальным макроэргическим фосфатом с АДФ, что дает еще 1 молекулу АТФ). Т.е., при полном окислении образовавшейся в гликолизе 1 молекулы НАДН и 1 молекулы пирувата получается 18 молекул АТФ, а 2-х – соответственно 36 молекул АТФ. С учетом того, что в процессе гликолиза образовались 2 молекулы АТФ, полный энергетический выход при окислении глюкозы до углекислого газа (CO2) и воды (H2O) в процессе клеточного дыхания, будет составлять 38 молекул АТФ.
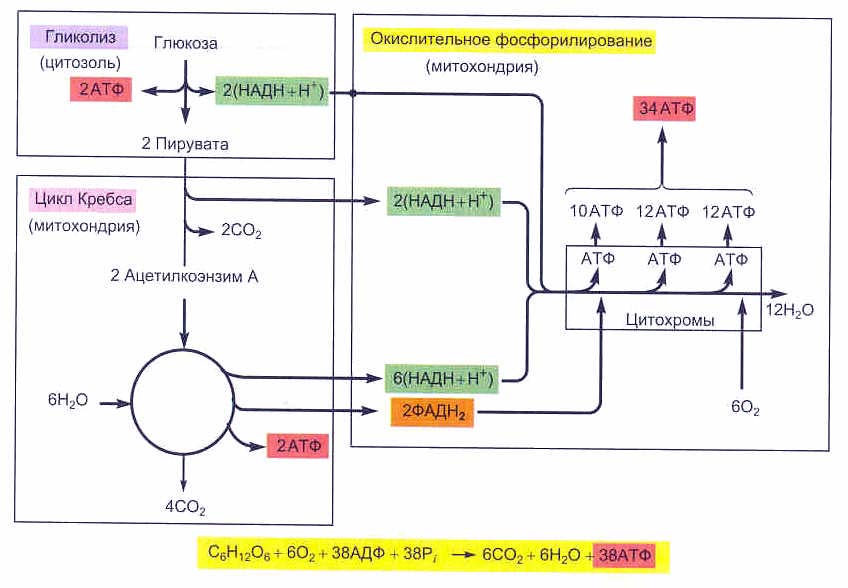
Полный энергетический выход окисления глюкозы до углекислого газа и воды в процессе клеточного дыхания составляет 38 молекул АТФ
Итоговое уравнение данного процесса будет выглядеть следующим образом:
С6H12O6 + 6О2 + 38АДФ + 38Фн —> 6CO2 + 6H2O + 38АТФ
Эффективность полного окисления глюкозы до углекислого газа и воды очень высока: от 55 до 70% освобождающейся энергии (в зависимости от конкретных условий) запасается в виде макроэргических связей в молекулах АТФ; остальная часть энергии рассеивается в виде тепла.
Таким образом, основным продуктом реакций энергетического обмена является АТФ.